
Vision
Think of your vision
神奈川大学 人間科学部 人間科学科
吉澤達也研究室
Think of your vision
私たちの研究目標は色情報がどのよな視覚的様相またはそのメカニズムに寄与しているのかを明らかにすることである.主要な研究テーマは以下に示す4つのであり,心理物理学的アプローチを含めた幾つかの手法を用いて研究している.
我々は,反対色チャンネルの時間的性質に興味を持ち,新規に開発したLEDを光源とした光学系を使って心理物理的実験を行った.その結果,2つの反対色チャンネルの間に時間的分解能のような時間的特性の違いがないことを見出した(Uchikawa & Yoshizawa, 1993; Yoshizawa & Uchikawa, 1997).さらに,2つの反対色チャンネルうち一方の反対色チャンネルのみを活動化させる高輝度の等輝度刺激に対する時間的特性で非常に興味深い新しい特性を発見した.この反対色チャンネルの時間的インパルス応答に関する従来の知識では,低域通過型フィルタのタイプとしてモデル化されている.
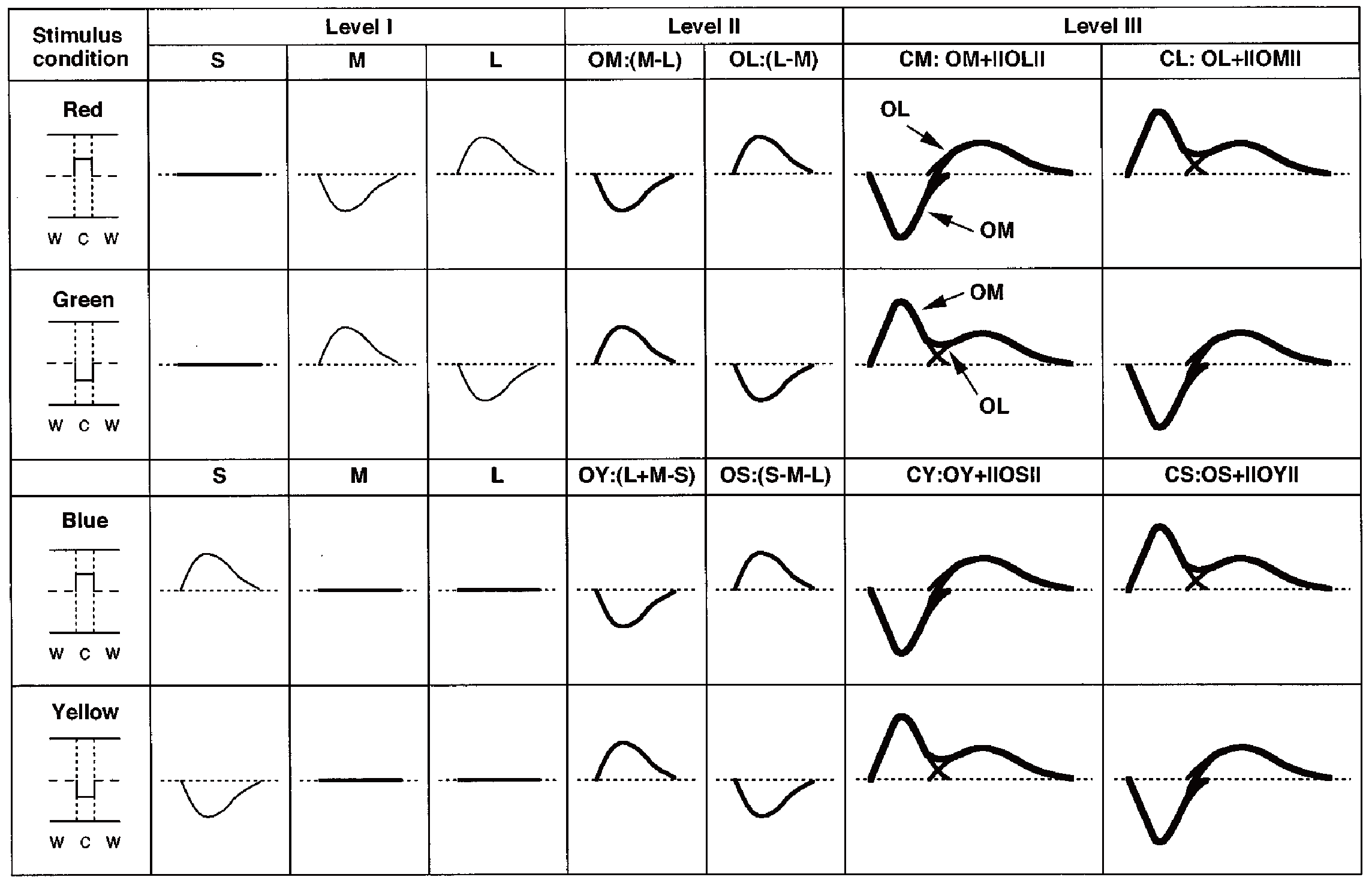 しかしながら,我々の結果は,時間的インパルス応答関数が,単純な低域通過型フィルタではなく,遅延を伴う2つの低域通過型フィルタ関数で構成されていることを示した(Yoshizawa & Uchikawa, 1997).反対色チャンネルの時間特性に関する進行中のプロジェクトは,反対色チャンネル全体の時間的特性を解明するために,MEGを用いた脳機能画像技術によって各皮質部位の時間的特性を調べている.
しかしながら,我々の結果は,時間的インパルス応答関数が,単純な低域通過型フィルタではなく,遅延を伴う2つの低域通過型フィルタ関数で構成されていることを示した(Yoshizawa & Uchikawa, 1997).反対色チャンネルの時間特性に関する進行中のプロジェクトは,反対色チャンネル全体の時間的特性を解明するために,MEGを用いた脳機能画像技術によって各皮質部位の時間的特性を調べている.
運動知覚における色情報の働きの仕組みについて精神物理学の研究を行った.従来,色情報は運動検出信号を生成するのに寄与しないと考えられてきた.これは,V1およびMTのような皮質における運動知覚検出機序が広範囲に渡ってフラットな分光感度を有しており,また,V4などの色覚機序が低い時間分解能を有するため,時空間の輝度変化がなく,色度変化のみによって定義される運動検出信号を生成することができないためである.しかし,近年いくつかの研究で,色情報が運動知覚のための信号をコード化することができることを報告しており,上記の従来の考え方について議論を引き起こしている
 それ以来,我々はこの問題を精神物理学的,生理学的,脳イメージングアプローチにより取り組んできた.我々は,逆位相の赤と緑の成分によって構成された正弦波グレーティング(1次色刺激)と色調コントラスト変調グレーティング(2次色刺激)の動きの方向弁別の性能を測定した.また,輝度グレーティングとコントラスト変調グレーティングの性能も測定した.その結果,我々は,一次色運動の性能は重畳された動的ルミナンスマスキングノイズの影響を受けやすいことから,一次色運動は輝度システムによって媒介されることを見出した.一方,2次色運動は真の反対色チャネルによって媒介されることを見出した(Yoshizawa et al., 2000; 2003; Mullen et al., 2003; Michna et al., 2007).
それ以来,我々はこの問題を精神物理学的,生理学的,脳イメージングアプローチにより取り組んできた.我々は,逆位相の赤と緑の成分によって構成された正弦波グレーティング(1次色刺激)と色調コントラスト変調グレーティング(2次色刺激)の動きの方向弁別の性能を測定した.また,輝度グレーティングとコントラスト変調グレーティングの性能も測定した.その結果,我々は,一次色運動の性能は重畳された動的ルミナンスマスキングノイズの影響を受けやすいことから,一次色運動は輝度システムによって媒介されることを見出した.一方,2次色運動は真の反対色チャネルによって媒介されることを見出した(Yoshizawa et al., 2000; 2003; Mullen et al., 2003; Michna et al., 2007).
この発見は,そのようなわずかな輝度変化で運動検出信号をどのようにコード化することができるのか,また,どのようなメカニズム特性が2次色運動を媒介しているのかという疑問を生じさせる.
最初の質問に対する仮説は,わずかな輝度信号を補償するために,色度変化と輝度変化の両方に基づいた運動検出信号を符号化できる神経細胞が存在する必要があるということである.2番目の質問に対する仮説は,まず,V4の神経細胞が順番に色度変化を分析し,次にMT細胞がV4から受信した情報を用いて運動検出信号を生成するという仮説である.
最初の仮説を確認するために,心理物理学的アプローチから生理学的アプローチへと研究方法を変更した.その結果,サルの電気生理学的取り組みにおいて,色度と輝度の両方に感度を持つV1細胞で,特にy / b反対色刺激に反応する神経細胞を見出しました.先行研究では,色度および輝度の変化(Johnson & Hawken,2001)の両方に感度を持つV1細胞が見出されたので,色,特にy / b変調および輝度変化の両方に感度を持つV1細胞が存在する可能性が予測された.しかし,まだ十分な生理学的データが得られておらず,これは進行中のプロジェクトである.
一方,2つ目の仮説を検証するために,MEGを用いて脳機能画像を用いて問題について研究してきた.こちらも,現在進行中のプロジェクトである.
1.M.L.Michna, T.Yoshizawa and K.T.Mullen (2007) Chromatic mechanisms mediating motion for S-cone isolating stimuli, Vision research, 47, 8, 1042-1054
2.T.Yoshizawa, K.T.Mullen, C.L.Baker Jr., Michael J. Hawken and Tetsuo Kawahara (2003) Contribution of chromatic information on motion perception and models of its mechanism, Japanese Journal of Visual Science, 24, 54-59 (in Japanese)
3.K.T.Mullen, T.Yoshizawa and C.L.Baker Jr. (2003) Luminance mechanisms mediate the motion of red-green isoluminant gratings: the role of temporal chromatic aberration, Vision Research, 43, 11, 1237-1249
4.T.Yoshizawa, K.T.Mullen and C.L.Baker Jr. (2003) Failure of signed chromatic apparent motion with luminance masking, Vision Research, 43, 7, 751-759
5.T.Yoshizawa (2002) Contribution of the number of Gabor micropatterns to dissociation of quasi-linear and nonlinear chromatic motion, The Journal of the Vision Society of Japan, 14, 57-68 (in Japanese)
6.T.Yoshizawa, K. T. Mullen and C. L. Baker Jr. (2000) Absence of a pure color linear motion mechanism in human vision, Vision Research, 40, 1993-2010
In third, I am interested in a recognition system of three-dimensional structure of visual objects. Human retina can get two-dimensional images, not three- dimensional ones. Therefore, our recognition system, in principle, has to produce information of three-dimensional structure of objects from the two-dimension retinal images. This ill-posed problem causes ambiguity of three-dimensional structure.
I investigated on this ambiguity in the structure-from-motion (SFM) stimulus. I found that relative luminance of moving dots of SFM to a background, which maybe be saliency of objects to a background, determined information of a depth order. That is, salient and/or high contrast dots appeared to be in front. This cue is important information to rule three-dimensional structure and robust to other parameters of the stimulus, such as a number of dots and a size of a stimulus. Furthermore, this featured stimulus with this saliency can induce three-dimensional structure of surrounding SFM stimulus that is ambiguous in depth order. This indicates the saliency has also a key role for contextual perception (Yoshizawa et al., 2006, 2007). Moreover, I have tackled to elucidate a mechanism of the biological motion (BM) perception.
There has been some debate as to the relative dominant cue to determine this perception: one is a form cue, and another is a motion cue. Most of the studies on this issue have tried to change parameters of the point-light BM stimulus and/or modify the stimulus in order to solve this problem. Because of changing or modifying the stimulus per se, different results were obtained on the base of different stimulus configurations. I therefore used a unique technique to extract crucial information in the BM perception. I superimposed a BM stimulus on the back-scroll illusion (BSI) background and changed a size and a location of the BSI background. The BSI is a phenomenon that for a movie so that a person makes a treadmill step against a counterphase grating background, we perceive that the counterphase grating moves toward the opposite direction of the walking person at the treadmill. This illusion was robustly observed with a point-light walker, as well as the case of a walking person in a movie (Fujimoto, 2003). If a motion cue is dominant and crucial to the BM perception, discrimination performance of the moving direction of a point-light walker should be changed by a size and a location of the BSI background. Whereas, if a form cue is more dominant than a motion cue, the performance should not be changed. It is because observers in our experiments could see a pint-light walker stimulus during the whole stimulus presentation time and could get form information. Our results showed that the performance was dependent of a size and a location of the BSI background. This indicates that a motion cue is dominant and crucial to the BM perception (Yoshizawa et al., 2009). Based on this finding, I would like to investigate each role of motion and form cues, its interaction and relationship between the cues in the BM perception mechanism to understand a mechanism for three-dimensional structure perception.
1.T.Yoshizawa, T.Kawahara and D.Kato (2009) On the mechanism of a movement direction in biological motion perception as inferred from the backscroll illusion, Perception (supplement), 38
2.T.Yoshizawa, M.Nakade, T.Kawahara and K.Maeda (2007) Effect of contextual cues on motion-surface labeling in structure from motion, Perception (supplement), 36
3.T.Yoshizawa, T.Kawahara and D.Nakayama (2006) Effect of contrast on motion surface labeling in 3D structure-from-motion, Perception (supplement), 35
I have been investigating on a contribution of color signal to binocular vision. As well as the issue of color and motion, it also has been a long controversial issue whether color signal contributes for the stereovision, especially to code binocular disparity information.
Previous studies by Simmons and Kingdom (1994, 1995, 1997, 2002) showed the contributions of color signal to binocular vision, whereas, for instance, results of Krauskopf and Forte (2002) study were negative to this contribution. Because these studies focused only the r/g opponent channel, but not the y/b channel, I tested on this issue of the y/b channel. It has been widely believed that the y/b opponent channel does not feed any signal to code luminance; therefore our approach was very suitable to solve the controversy. I found that the y/b signal could contribute to code the binocular disparity signal, even though the s-cones are sparsely distributed in fovea (Yoshizawa et al, 2008). I have been still investigating on the residual problems of the issue in psychophysical approaches.
1.T.Yoshizawa, H.Kobayashi, T.Kawahara and K.Maeda (2008) Interaction between y/b chromatic and luminance mechanisms in disparity detection, Perception (supplement), 37
2.T.Yoshizawa, H.Kobayashi, T.Kawahara (2008) S-cone channel contributes to the depth perception, Asian-Pacific Conference on Vision, 67